
Что такое миграция рыб
Миграция рыб
От лосося, который может преодолевать водопады одним мощным прыжком, до тунца, обтекаемого, как летящая пуля, многие рыбы являются мастерами далеких путешествий.
Мигрируют и пресноводные, и морские рыбы. Их путешествия к местам размножения могут быть разделены на четыре основные категории, базирующиеся на особенностях биологии: из пресной воды в морскую, из морской воды в пресную, из одного морского региона в другой и из одного пресноводного района в другой.

Как правило, большинство видов угрей обитают в морях, но виды семейства пресноводные угри (Anguillidae), к которому отнесен и европейский речной угорь (Anguilla anguilla). растут и развиваются в пресных водоемах, а затем мигрируют в море для размножения. Эти путешествия носят специальное название катадромных миграций.
Европейский угорь начинает свою жизнь в Северной Атлантике, в Саргассовом море. Здесь из икринок выводятся десятимиллиметровые личинки, называемые лептоцефалидами. Раньше ученые считали этих личинок самостоятельным видом животных, не имеющим никакого отношения к взрослому угрю. Когда личинки подрастают, они начинают свое путешествие, пассивно дрейфуя из Саргассова моря. Через Атлантику их несет течение Гольфстрим, и через три года выросшие личинки достигают берегов Европы. Входя в устья рек и попадая в пресную воду, личинки превращаются в молодого угря или в так называемую стадию «стеклянного угря». Стеклянные угри большими стаями движутся против речного течения в поисках мест обитании, завершая эту фазу миграции, и все путешествие, в ходе которого они преодолели путь длиной более 5000 км.
Угри остаются в реке на несколько лет, до тех пор, пока не превратятся в совершенно созревших взрослых рыб. Удивительно, но взрослых европейских угрей, путешествующих вниз по течению реки, для того чтобы снова достичь моря, мало кто видел. Их североамериканские двойники, такие же взрослые угри, регулярно встречаются спускающимися вниз по течению. Когда они достигают Атлантического побережья Северной Америки, то начинают финальную фазу своей миграции. Рыбы направляются в Саргассово море — традиционное место размножения, тем самым завершая свой жизенный цикл.
На протяжении своей жизни и для навигации во время миграции угри используют многие ориентиры. Они не только очень чувствительны к запахам, но с готовностью реагируют на изменения в движении воды, сейсмическую активность и на самые слабые электрические поля, генерируемые потоками воды. В миграциях личинкам угря помогают океанические течения — североамериканские угри движутся к северу вдоль восточною побережья, а их европейские сородичи через Атлантику вместе с Гольфстримом.
Некоторые ученые выдвигают гипотезу о том, что североамериканский и европейский угри являются одним видом. Они также предполагают, что часть европейских угрей погибает, завершив свой миграционный цикл, а североамериканская форма пополняет численность обеих форм в Саргассовом море. Критики этой теории обращают внимание на то, что у европейского и американского угрей в скелете разное количество позвонков. Причем количество позвонков у этих рыб может изменяться под влиянием окружающей среды. Угри, выращенные при более высокой температурю воды, имеют больше позвонков.

Икрометание лосося
Миграции рыб, которые для икрометания из открытого океана идут в пресноводные реки, поднимаясь против течения, называются анадромными. Наиболее известным представителем анадромных рыб является лосось. Эта рыба отмечалась за медленное и трудное путешествие, которое она совершает, возвращаясь из океана, где проводит большую часть жизни, к местам своего рождения в маленьких пресноводных речках и озерах.
Атлантический лосось (Salmo salar) начинает свою жизнь весной в одном из бурных горных потоков Норвегии или Шотландии, где он появляется из икринок. В реке лососи нередко проводят более четырех лет, прежде чем начинает обратную фазу своей марафонской миграции. В это время молодые лососи, готовые к миграции, путешествуя вниз по реке к океану, получают название серебрянка, или смолт. Все это время физиология серебрянки постепенно меняется, приспосабливаясь к обитанию в соленой морской воде.
Путешествуя и взрослея в океане, лососи проводят там более четырех лет, а затем начинают обратное путешествие домой. Став взрослыми, рыбы самостоятельно отыскивают дорогу из океана в ту самую реку, в которой они появились на свет много лет назад.
Необыкновенная способность лосося находить реку, в которой он родился, связана с высокоразвитым чувством обоняния, способным различать воду разных рек по запаху.
Выживание сильнейших
Возвращаясь к своему нерестилищу, лососи плывут против течения реки и часто бывают вынуждены на своем долгом и изнурительном пути преодолевать пороги, водопады и другие естественные преграды, а также препятствия, созданные человеком. Только самые приспособленные и самые сильные способны совершить это путешествие. Только они достигают своей реки и оставляют потомство. Нерестовая миграция — важный процесс естественного отбора, в ходе которого только лучшие из лучших рыб дойдут до цели, для того чтобы передать свои ценнейшие гены следующему поколению.
Более того, у атлантического лосося часть взрослых рыб после размножения пускается в обратный путь, скатывается по рекам и возвращается в океан, где изнуренные рыбы постепенно восстанавливают свои силы и живут еще несколько лет, а их тихоокеанские собратья, лососи (Oncorhynchus spp.), гибнут после икрометания.
За время своего путешествия лососи преодолевают громадные расстояния, достигая севера Гренландии и побережья Норвегии и иногда покрывая расстояние более 5600 км.

Pacific bluefin tuna (Thunnus thynnus) schooling, Mexico.
Путешествующие тунцы
Хотя тунцы никогда не покидают свой океан и считаются океанодромными рыбами, они нередко предпринимают миграции более дальние, чем даже такие известные путешественники, как лососи и угри. После икрометания, которое проходит поздней весной и ранним летом, в морях Флориды и Багамских островов, а в Средиземном море — в середине лета, голубой тунец (Thunnus thynnus) иногда предпринимает дальние миграции к северу. Эти путешествия тунцы предпринимают в поисках своей обычной добычи — более мелких рыб, таких, как сельдь и скумбрия.
В целях изучения миграций гунна, уточнения используемых ими морских путей и пройденных расстояний ученые пометили множество гуннов индивидуальными метками, которые называются дротик и крепятся к телу рыбы. Эти индивидуальные метки содержат детальную информацию, касающуюся места мечения рыбы, и те, кто ловит помеченных тунцов, возвращают метки обратно ученым.
Анализ встреч помеченных рыб показал, что одна из особей, помеченных в 1958 году, в мексиканском Калифорнийском заливе, была поймана пятью годами позже в 483 км южнее Токио (Япония). Это означает, что минимальная (то есть по прямой линии) дистанция, которую он проплыл за эти годы, составляет 9335 км. Конечно, рыба преодолела намного большее расстояние, потому что ни одна рыба не может на такой огромной дистанции плыть строго но прямой.
Из озера в реку и обратно
Среди рыб, мигрирующих в пресных водах, есть такие виды, как карпы, усачи, гольяны, сомы-кошки, а также и громадный североамериканский панцирник.
Смысл пресноводной миграции состоит в том, что рыба, избегая многочисленных хищников, для размножения перемещается из глубоких, медленно текущих рек или озер, к быстротекущим мелководным притокам. Эти специфические места размножения мигранты активно ищут и легко узнают, ориентируясь в пути по запахам.
Отметав икру, рыбы возвращаются обратно к местам постоянного обитания, где завершают свой жизненный путь.
МИГРАЦИИ РЫБ
Миграции рыб, как и других животных,— это массовые, обычно активные, но иногда и пассивные перемещения из одного местообитания в другое. Рыбы перемещаются туда, где они находят условия, необходимые организму на той фазе жизненного цикла, которая наступит к концу миграции.
Миграции, как и другие видовые свойства, имеют приспособительное значение, обеспечивая благоприятные условия существования и воспроизводства популяции вида. Миграция представляет собой звено жизненного цикла, неразрывно связанное как с предыдущими, так и с последующими звеньями.
Миграционный цикл обычно состоит из размножения или нагула к местам зимовки.
Схематически миграционный цикл рыбы, да и многих других животных, может быть представлен в следующем виде (рис. 109). По характеру совершаемых миграций рыбы подразделяются на мигрирующих и оседлых. Лишь сравнительно немногие рыбы постоянно живут на одном и том же месте, не совершая закономерных более или менее длительных перемещений. К таким
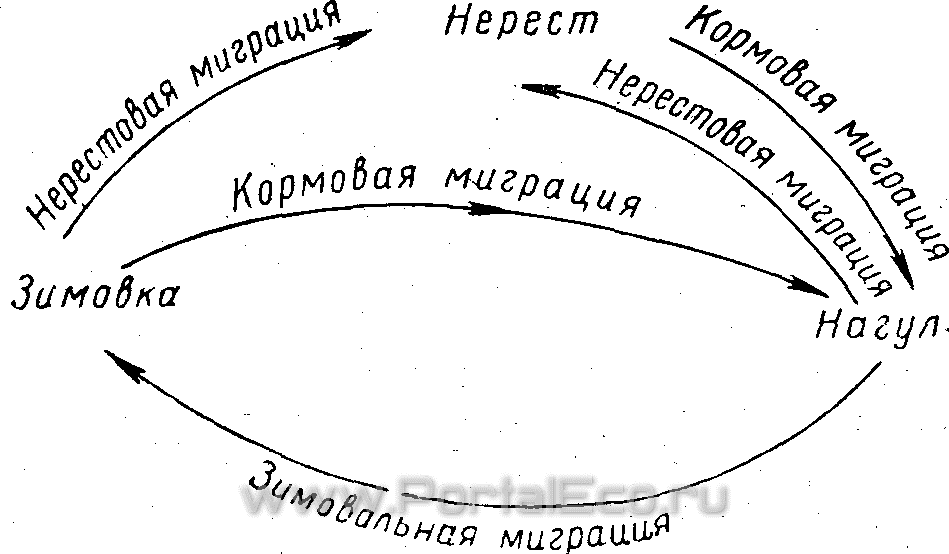
Схема миграционного цикла различных рыб
рыбам относятся, например, некоторые бычки семейства Gobiidae, многие коралловые рыбы семейства Pomacentridae, Siganidae, Apogonidae и др. У большинства рыб миграции составляют необходимое звено их годового цикла жизни.
Не у всех рыб миграции выражены одинаково. У одних (как, например, многие сиги) имеются только нерестовая и нагульная миграции, а зимовальная отсутствует. У других рыб, места размножения которых совпадают с местами нагула, как, например, у многих пескарей, имеется лишь зимовальная миграция, связанная с переходом по окончании периода нагула на более глубокие участки русла реки.
У одних видов миграции совершают только взрослые рыбы, достигшие половой зрелости, а молодь, скатившись от нерестилищ к местам нагула, живет здесь, не совершая значительных перемещений, как это имеет место, например, у многих лососей. У других рыб и молодь совершает столь же протяженные миграции, как и взрослая рыба. Это мы наблюдаем, например, у бражниковской сельди — Caspiatosa brashnikovi (Borodin), у которой и молодь и взрослые рыбы идут с мест зимовки из Южного Каспия в Северный Каспий, где располагаются места размножения и нагула этой рыбы.
Первый тип миграционного цикла, свойственный многим лососевым и другим видам рыб, имеет место в том случае, когда станция размножения и станция нагула находятся в разных местах, а станция зимовки и нагула совпадают.
Второй тип миграционного цикла характерен для тех рыб, у которых совпадают места размножения и нагула.
Поскольку миграция — это звено жизненного цикла, неразрывно связанное с предыдущими и последующими звеньями, то естественно, что рыба начинает миграцию достигнув определенного состояния, т. е. будучи подготовлена к ней.
Так, у большинства рыб начало нерестовой миграции связанно обычно с достижением определенной стадии зрелости половых продуктов и проявлением определенной гормональной активности железами внутренней секреции. При этом у рыбы меняется характер реакции на внешние воздействия, т. е. появляется новый натуральный раздражитель, являющийся сигналом для начала миграции.
Начало зимовальной миграции у большинства рыб связано с достижением определенной упитанности и жирности, обеспечивающих успешную зимовку рыбы. Подготовленный к зимовке аральский лещ имеет коэффициент упитанности около 3,0 (по Фультону) и высокий процент жира в мясе. Подготовленная к зимовальной миграции азовская хамса имеет коэффициент упитанности в среднем —1,00. Подготовленность к миграции еще не означает, что рыба сразу же начнет мигрировать. Миграция начинается лишь при определенных условиях. Худой лещ и худая хамса не начинают зимовальной миграции даже при резком снижении температуры воды. Но для подготовленных к зимовке леща и хамсы резкий перепад температуры становится натуральным раздражителем—сигналом, вызывающим начало зимовальной миграции. Так, по данным Г. Е. Шульмана (1959), хамса, имеющая до 14% жира, не мигрирует при любом перепаде температуры. При жирности 14—17% для начала миграции необходим перепад температуры 9—14°, и ход бывает растянутый и недружный. При достижении жирности 22% хамса начинает мигрировать при любом перепаде температуры, и ход бывает дружный.
Таким образом, начало миграции зависит как от состояния самой рыбы, так и от изменения окружающих условий. Срок начала миграции вырабатывается как приспособление, обеспечивающее благоприятные условия и для самой рыбы (в случае зимовальной и нагульной миграции) и для ее будущего потомства в случае нерестовой миграции.
Нерестовая, нагульная и зимовальная миграции находятся во взаимной связи, .они подготовляются предыдущими звеньями жизненного цикла и сами обеспечивают дальнейшую жизнь рыбы. Как отмечалось, переход в миграционное состояние всегда связан с определенным состоянием рыбы — ее упитанностью, жирностью, развитием половых желез и т. д.
Наряду с миграциями, являющимися звенрм жизненного цикла рыбы, многие рыбы совершают массовые перемещения, которые происходят при разном биологическом состоянии рыбы и носят чаще всего защитный характер. Примерами подобного рода перемещений могут служить отходы рыбы из прибрежной зоны во время волнения, отход рыбы из озер и заливов во время падения уровня воды в реках и др.
Большинство рыб совершают миграции, активно двигаясь в том направлении, в котором лежит их миграционный путь, и, следовательно, затрачивая на миграции энергетические вещества, накопленные в их теле. Но у многих рыб наряду с активными имеют место и пассивные миграции, когда рыба или наиболее часто ее пелагическая икра пассивно переносятся на весьма значительные расстояния. При этом организм не расходует энергии на движение, ибо оно осуществляется вместе с перемещением водных масс — течениями. Во многих случаях часть миграционного пути преодолевается путем пассивной миграции, а часть — активно, как это имеет место, например, у личинок обыкновенного угря при их миграциях от берегов Центральной Америки к берегам Европы. Сначала этот путь совершается пассивно вместе с водами Атлантического течения, а при приближении к берегам угорьки начинают уже активно мигрировать в реки.
Каковы же те причины, которые привели у различных групп рыб к выработке столь сложного биологического явления, какое представляет миграция?
Несомненно, что миграция есть приспособление, обеспечивающее существование вида. Допустим, например, что все лососи, которые входят для икрометания в ту или иную реку, остались бы в ней на кормежку, а не ушли бы для нагула в море — несомненно, что очень скоро кормовые ресурсы реки были бы исчерпаны, и поголовье стада должно было бы сократиться. Естественно, что самки, расходующие большое количество энергии на развитие икры и обычно бывающие у проходных рыб больших размеров, чем самцы, для обеспечения как собственного существования, так и развития икры, больше чем самцы нуждаются в усиленном питании. Это питание в достаточном количестве они могут получить только в море. Действительно, в природе мы наблюдаем, что у многих рыб самцы или часть самцов постоянно живут в реке, а самки ведут проходной образ жизни. Это явление зарегистрировано у лосося, усача и некоторых других рыб.
Но у нас естественно, возникает вопрос, почему тогда проходные рыбы, находящие в море более благоприятные условия для питания, не переселились полностью в море, как это произошло с рядом ныне живущих в море генеративно-пресноводных рыб, а сохранили свои места размножения в пресной воде?
Как известно, большинство морских рыб, откладывающих донную икру, ее охраняют. В противном случае икру очень быстро нацело съедали бы разные хищники. Следовательно, при переносе нерестилищ в море проходным рыбам пришлось бы охранять икру, так как иначе она была бы съедена. Почти все проходные рыбы (кроме миног и сельдей) как крупные животные, имеющие значительный охотничий район и совершающие большие кормовые перемещения, вынуждены были бы при охране икры в течение длительного инкубационного периода часто голодать, а то и гибнуть от голода до окончания периода охраны икры, оставляя ее беззащитной. В реках же и без активной охраны развивающаяся Икра подвергается значительно меньшим опасностям, чем в море. Закапывать в море икру в грунт, как это делают лососевые в реках, очевидно нельзя из-за неблагоприятного кислородного режима грунтов значительных глубин и постоянной подвижности гальки прибрежной зоны.
Таким образом, переход на кормежку в море позволяет виду значительно увеличить свое поголовье за счет часто огромных кормовых ресурсов морских водоемов, а размножение в реке обеспечивает большее выживание молоди. Прирост поголовья стада ограничивается, однако, и величиной нерестовых площадей. Как указывает И. И. Кузнецов (1928), с увеличением концентрации гнезд на нерестилищах лососевых (свыше одного гнезда на 2 м2), резко возрастает процент гибнущей в гнездах икры, что лимитирует величину поголовья стада. Площадь, пригодная для нерестилищ в реках, далеко не безгранична, а часто бывает и очень незначительной.
Следовательно, у проходных генеративно-пресноводных рыб основной причиной возникновения миграционного цикла является недостаточная обеспеченность кормом в реках.
Миграции рыб
Условия существования рыбы не являются чем-то неизменным и могут меняться самым различным образом. Эти изменения окружающей обстановки в связи с изменениями, происходящими в самом животном организме в зависимости от возраста, созревания половых продуктов и т. п., и являются причиной того явления, которое носит название миграций рыб и имеет огромное практическое значение.
Миграции можно разделить на пассивные, вызываемые, например, течениями, и на активные, когда животные переселяются, отыскивая для себя лучшие, более подходящие условия. Самое большое значение имеют те миграции, которые связаны с размножением, с нерестом (нерестовые), и миграции молоди (личиночные). Ho, кроме этих миграций, существуют еще сезонные периодические миграции, куда относятся переселения рыбы на места зимовки, из глубин па поверхность и на мелкие места, а также короткие или долгие странствования вверх и вниз но реке, — одним словом, миграции, обусловленные изменениями температуры, отыскиванием пищи и т. д.
Примитивные рыбы или являются пресноводными или мигрируют в пресную воду для размножения (Lampetra, Chondrostei). Акулы имеют демерсальные прикрепляющиеся яйца, и их палеозойские предки были или пресноводными или посещали пресные воды.
Демерсальные яйца имеются у многих Clupeidae, у лососей (Salmonidae), у осетровых (Acipenseridae). Пелагические яйца имеются главным образом у морских рыб. Развитие пелагических яиц сделало возможными жизнь и икрометание в открытом море.
Личинки (правильнее — молодь) рыб развились также в разных направлениях. Одни, как личинки тихоокеанского лосося, плывут сотни миль вниз по реке, чтобы достичь моря; другие, как личинки лосося атлантического, опускаются на дно, прячутся под камни, чтобы не быть унесенными течением, или зарываются для этого в песок, прикрепляются и т. д. Третьи не сопротивляются течению, плывут пассивно и уносятся иногда очень далеко от места вывода из яиц. Таким образом, и личинки могут быть подразделены на пелагические, переносимые течениями, и на погружающиеся.
Там, где течений нет или где усилия рыбы бороться с ними увенчиваются успехом, там места икрометания и места кормления молоди почти или вполне совпадают. В большинстве же случаев такое совпадение бывает редко, оба эти места лежат далеко друг от друга, и рыбам приходится совершать большие переселения.
Изучение миграций даже главнейших промысловых рыб требует громадной работы в различных направлениях. Прежде всего необходимо уметь отличить данный вид на каждой стадии его развития, отличить яйцо, личинку, молодь одного вида от другого, для чего собираются и изучаются обширные коллекции. Для определения рас, которые па основании обычных морфологических методов неразличимы, выработаны особые математические методы, о которых говорилось вышe.
Изучение физических и химических свойств морской воды, ее температуры и течении также оказывает помощь в изучении странствования рыб, в особенности важно оно для понимания путей странствования более молодых классов рыбы, которые плавают пассивно.
Самое изучение миграций может производиться различными методами. Весьма точный и верный из них — это «мечение» рыбы, когда том или иным способом к пойманной рыбе прикрепляют металлическое кольцо или пластинку с обозначенном места и числа, когда рыба мечена, и затем рыбу выпускают. Метить возможно лишь более крупные экземпляры, и это было сделано в большом объеме и с превосходными результатами с камбалами у берегов Исландии и с лососевыми в Аляске.
В некоторых случаях прекрасным методом изучения миграции являются ловля планктонных яиц, личинок, молоди и рыб на разных стадиях их развития и сопоставление всех этих данных. Таким методом были изучены в 1901 г. нерестилища трески в северной Норвегии. Было сделано большое количество топей во всех направлениях между банками и добыто большое количество яиц. Считая эти яйца после каждой топи, нашли, что топи продолжительностью в 5 минут, сделанная при помощи маленькой шелковой сетки в 1 м в диаметре, давала тысячи яиц, плававших над этими банками на глубине 60—80 м, где рыба, метала икру. Раз было найдено нерестилище, оно было использовано и дало не менее полутора миллиона экземпляров трески. Этим же методом была выяснена история странствования угрей.
Точно так же можно определять пути странствования, отмечая различия в величине рыб разных пунктов.
Переселения, связанные с размножением, с отыскиванием места нереста, бывают различного типа.
Крайние типы представляют миграции угря и лосося. Первый, достигая зрелости, скатывается вниз по рекам и плывет против Гольфштрема, чтобы достичь места икрометания среди океана; второй входит по достижении половой зрелости в реки и идет для икрометания вверх по последним. Первый тип миграции носит название катадромальной миграции, второй — анадромальной миграции.
Явления миграций станут нам более ясными из рассмотрения отдельных типичных случаев катадромальной и анадромальной миграций.
Катадромальная миграция угрей (Anguilla anguilla). Вхождение молодых угрей в реки весной и скатывание в море «серебристых угрей» осенью были известны давно. Лишь недавно было доказано, что так называемые тонкоголовые (Leptocephali), считавшиеся за отдельный род рыб со многими видами, являются в действительности личинками угрей, в которых они превращаются. Местами размножения угрей предполагали глубины Атлантического океана.
В настоящее время работами норвежских и датских ученых тайна угря разгадана, и миграции угрей представляются в таком свете.
Существуют два вида угрей: европейский угорь — Anguilla anguilla и американский — Auguillarostrata, различающиеся между собой по количеству позвонков: 114 у европейского и 107 у американского. В конце лета почти половозрелые угри спускаются из рек и озер в моря — Балтийское, Северное, Средиземное, а отсюда в океан, к местам икрометания (рис. 206). Последнее удалось установить лишь совсем недавно. Места икрометания находятся для обоих видов близко друг от друга, частью перекрываясь, в так называемом Саргассовом море, в западной части Атлантического океана. Здесь яйца откладываются на большой глубине (800—900 м). Выметав икру, угри, видимо, погибают, так как к концу нерестования в океане уже не видно взрослых угрей. В конце зимы или начале весны из икры выводятся молодые угри в виде листообразных личинок, называемых тонкоголовыми — Leptocepbali, Вырастая, они медленно поднимаются в верхние слои воды. Когда они поднимутся, то дальнейшая история жизни протекает неодинаково у американского и у европейского угря. Американские Leptoсу растут гораздо быстрее и уже в течение одного лета дорастают в среднем до 6—6,6 см, достигая в это время берегов своей родины. Зимой происходит метаморфоз в прозрачных «стеклянных» угрей, на вторую весну — вхождение в американские реки.
Миграция рыб
Рыбы путешествуют намного больше и охотнее, чем кажется людям. Общеизвестны странствия лосося и угря, области обитания которых располагаются за тысячи километров от мест нереста. Но знаете ли вы, что окуни и подусты отправляются к местам нереста, расположенным на расстоянии до 400 км?
Рыбы всех видов в течение года или даже дня перемещаются в различные участки водоема. Эти зоны используются ими как места кормления, нереста и зимовки. К тому же рыбы совершают миграции во время половодья, сезонные, при расселении, а также компенсационные. А на самых ранних стадиях жизни происходят дрейфы личинок и молоди.
Нерестовые миграции могут быть либо только внутри пресноводных водоемов, либо имеет место смена пресной воды на соленую. Лососи, кумжи и рыбы других видов, которые переходят для нереста из моря в пресную воду, называются анадромными. Противоположным путем идут так называемые катадромные рыбы, к которым относится угорь. Они плывут для размножения из рек и озер в океан. Рыбы, которые предпринимают нерестовые миграции внутри пресных вод, называются потамодромными.
Мигрируют и мелкие рыбы
Наиболее доступный метод получения информации о миграциях рыб – подсчет их в местах подъема в реки. Правда, такие исследования дают мало информации о дистанциях миграции, однако они показывают, какие рыбы и каких размеров склонны к миграции. Так было доказано, что и мелкие речные обитатели, такие как гольяны и красноперки, активно меняют жизненное пространство. Этих рыб долгое время считали немигрирующими. Интересен также и тот факт, что большинство поднимающихся в реки рыб являются неполовозрелыми.
Это показывает, что у рыб многих видов молодь тоже склонна к миграции. У форелей, хариусов, усачей, подустов и многих других речных рыб первые миграции происходят в личиночной стадии. Эта пассивная миграция, называемая еще личиночным дрейфом, служит для того, чтобы после нереста как можно быстрее заселить большие пространства водоема. В ходе научных исследований было доказано, что дрейф личинок – важнейший фактор при заселении вновь возникающих водных рукавов. Тем самым личиночный дрейф может быть интерпретирован как миграция распространения. Эта форма миграции служит для освоения новых жизненных пространств и выравнивания популяций различного размера. Заселение вновь созданных водоемов, например при расчистке старицы реки, происходит очень быстро. Максимальное число видов рыб наблюдается в водоеме уже через несколько лет. При высокой воде рыбы вынуждены уходить с насиженных мест в зоны с более спокойным течением. В большинстве случаев они покидают главный поток и переходят в притоки или в зоны с медленно текущей водой. При экстремальных половодьях заселяются и затопляемые зоны. Если вблизи водоема таких мест нет, многие рыбы скатываются вниз по течению. В таких случаях после спада воды происходят компенсационные миграции в безрыбные области.
Сезонные миграции
Под сезонными миграциями понимают реакцию рыб на обусловленные временем года изменения условий окружающей среды. Например, ручьевые форели осенью и ранней зимой после закончившегося нереста мигрируют в расположенные ниже по реке зоны с глубокими заводями и слабым течением. Тем самым они избегают уменьшения жизненного пространства при низкой воде и уходят от опасности промерзания мелководных участков водоема.

Весной, после таяния снега, эти рыбы вновь поднимаются вверх по реке и заселяют отдаленные родниковые ручьи. Так они оптимально используют имеющиеся в их распоряжении места в любое время года. Сезонные миграции могут быть обусловлены и питанием. Например, хищные рыбы следуют за меняющимися в течение года местами стоянки кормовых рыб. Особыми формами миграции, не попадающими под вышеупомянутую классификацию, являются суточные вертикальные миграции.
Рыбы некоторых видов в стоячих водоемах следуют за добычей в течение дня с глубины 60 м до водной поверхности. Например, сиги следуют за зоопланктоном, поднимающимся под защитой ночной темноты из глубины к поверхности воды, поэтому не удивляйтесь, если вечером неожиданно сиг схватит червя на вашем крючке.













